Cynops pyrrhogaster (Boie, 1826)
両生綱 > 有尾目 > イモリ科 > イモリ属 > アカハライモリ
 成体メス 岐阜県恵那市
成体メス 岐阜県恵那市




[分布]
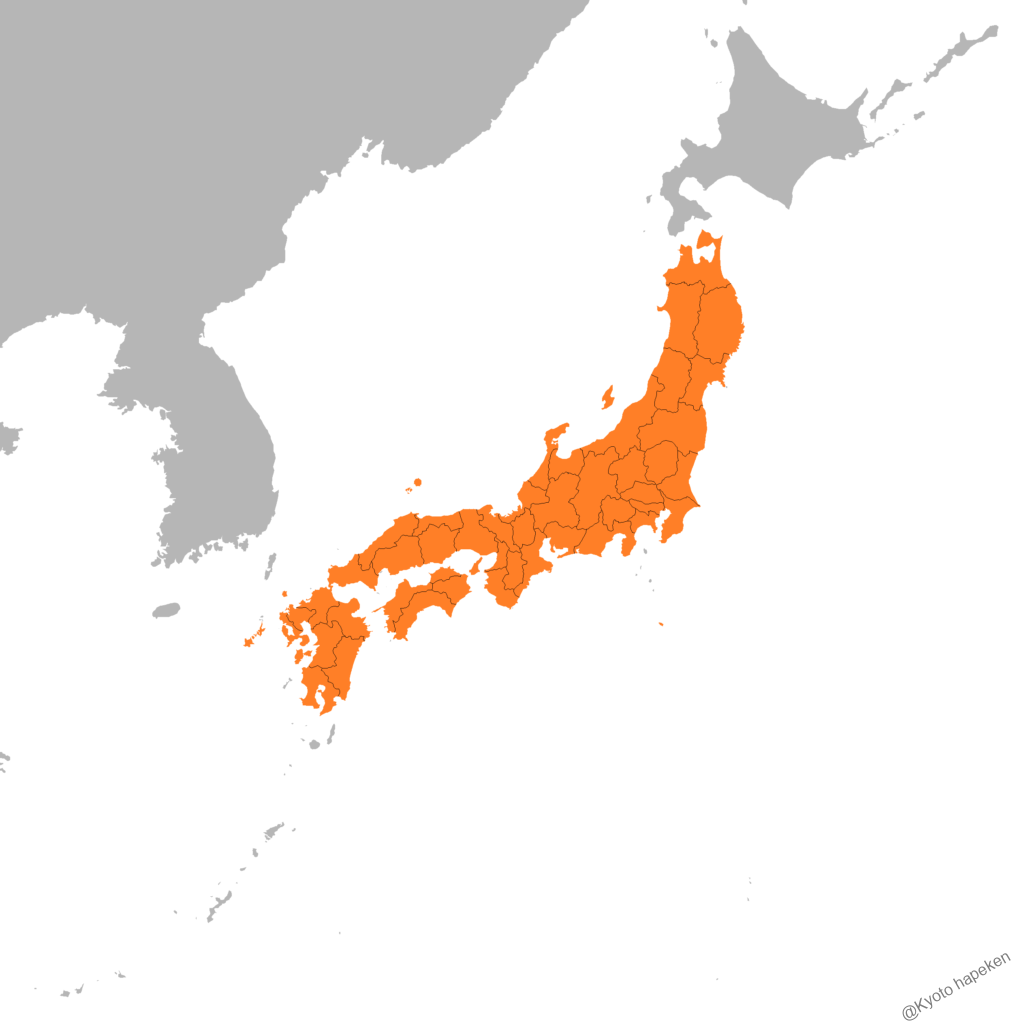
- 本州 四国 九州 佐渡島 壱岐 隠岐 五島列島 八丈島(四国からの移入) [14][22]
[生息環境]
- 止水かそれに近い環境の水中 幼体は林床など
[食性]
成体はオタマジャクシ、イトミミズ、ユスリカの幼虫、ハエ亜目の成虫、トンボの死体、魚卵など[3][18]の他、同種の卵や幼生も捕食する[3]。
幼体はトビムシやダニなど陸生の小動物を食べる[8]。
[捕食]
- ニホンマムシ、カラス、ゴイサギ等に補食された例がある[2]。
- 外敵に襲われた際には逃走するか立ち止まって身をそらし(スズガエル反射:unken reflex)腹側の赤色を見せて有毒であることをアピールする。水温が高いときは機敏に動けるためかより逃走を選択しやすい[10]。
- 島嶼部に生息するイモリの腹面は本土産のイモリに比べ赤色の面積が大きくより派手である傾向にあり、襲われた際に逃走せず身をそらすなどして腹面の赤色をアピールする頻度も高い。警戒色によるアピールは色覚に優れる鳥類に対しより有効な防御手段であるが哺乳類の捕食者(大部分が色盲)にはあまり効き目が無いと思われることから、島嶼部のイモリがより派手な腹面とそれを誇示しやすい性質を持つのは、一般に島には哺乳類の捕食者が少なく、相対的に鳥類に襲われる確率が高いためと考えられている[9]。
[繁殖]
繁殖期のオスは尻尾や頸部及び体側に薄紫の婚姻色を呈するが、渥美種族と呼称される個体群のオスには婚姻色が見られない[16]
交配行動は以下のように進行する[2][4][11][13]
➀識別
繁殖の準備が出来ているメスは総排出孔からアイモリンというフェロモンを出していて、オスは吻端を別個体の体側にあてることでフェロモンの有無を調べ、相手が求愛の対象であるか識別する。
②求愛
相手がメスであることを確かめるとメスの前で横になって進行を阻害し、尾を振り、ソデフリンを水流に乗せて送ってメスに求愛をし始める。
③反応
オスは大抵の場合振られてしまうが、求愛を受け入れたメスはオスの頸部や胴体を吻端でつつくなどして了承のサインを出す。
④従行
サインを受けたオスはメスから離れ、尻尾をくねらせながら歩き出す。メスはその後ろをついて歩く。この際メスの吻端は常にオスの尻尾に触れている。
⑤精子塊の放出
オスが停止し、尻尾をくねらせつつ持ち上げ、粘着性の高い精子塊を一つ落とす。
⑥受け入れ
オスは再び前進を始め、メスは後を追う。メスが精子塊の上を通過する際に総排出孔から精包を吸収する。精子塊の放出と吸収は普通一回だけでは終わらず、④~⑥の流れは2,3回繰り返され、従って一回の交配に付きオスは2,3個の精子塊を放出することになるが、メスがその全てを吸収できるとは限らず、むしろ1つか2つ置き去りにするか、足や腹面に付着させてしまったりする事が多い。
また一連の交配行動には地域差がある。例として
・②の求愛で篠山、渥美、中間種族のイモリはメスの頭に後肢を乗せる(渥美のイモリはこの行動を取らないことも多い。)
・関東種族のイモリのメスは③の反応の際にはオスの頸部を、④の従行の際には尾部をよく噛む
等がある。こうした違いのためか異なる地域種族間では交配が上手くいかないことが多い。とはいえ野生下で地域種族間の遺伝的交流が全くないわけでは無く、異なる種族同士の間にも交雑帯が存在する[17][21]。
繁殖期は4月から7月。しかし10月ごろから冬眠までの間に婚姻色を呈しメスと交尾するオスもいるらしく、岡山県上斎原のイモリが冬眠明け以降交尾をしていないにもかかわらず産卵し幼生が産まれた例が観察されている[1]。

[卵]
- 卵は球形で一つずつ水草などに産みつけられる。一回の産卵数は40個以内で、長期にわたって産卵し合計100から400の卵を産む[2]。
[幼生]
- 3対の鰓を持ち、完全な水棲。孵化直後はカエルの幼生であるオタマジャクシと同じく手足を持たないが、孵化して5日ほどの間は眼と鰓の間にバランサーという棒状の突起が一対存在しこれにより体を支える(バランサーはやがて脱落してしまう)。その後カエルとは逆に前肢、後肢の順に足が生えてくる。孵化後3~4ヶ月で鰓は消え、変態し幼体となる。体長はふ化直後で10~12mm 変態前で50mmほど[18]。
[幼体]
- 変態して幼体になると上陸し、成体になり水中生活に移行するまでの間は陸上で生活する。幼体の期間は低地で三年ほど、高地ではそれ以上[3]。飼育下では上陸の時期に水位を下げるか陸地を設けるかしないと幼体は溺死してしまう。
[成体]
- 夜行性だが日中にも活動し、繁殖期には特に日中の活性が上がる。また飼育下では日中でも問題なく餌を食べる。成体となり水場に戻ったイモリはかなり水棲傾向が強いが雨の日などに陸に上がることもあり、また真夏などに水温が上昇した際には水場周辺の林地に散ってしまうらしい[15]。
- 8月と9月に最も近い水場から1km近く離れた山の頂上付近(陸上)で成体が見つかったという報告もある[5]。また冬眠は陸でも水中でも行う。
執筆者:市岡幸歩
引用・参考文献
- Akiyama, S., Iwao, Y., & Miura, I. (2011). Evidence for true fall-mating in Japanese newt Cynops pyrrhogaster. Zoological science, 28(10), 758-764.
- 日高敏隆. 1996. 日本動物大百科第五巻. 平凡社, 東京. p.24-27.
- Ihara, S. 2014. Food Habits of the Adult Japanese Newt Cynops pyrrhogaster (Amphibia: Salamandridae) in the Sub-Alpine Yachidaira High Moor, East—Central Honshu, Japan
- Kikumura, S., F. Toyoda, Y. Ohmiya, K. Matsuda, S. Tanaka, and H. Hayashi. 1995. Sodefrin: A female-attracting peptide pheromone in newt cloacal glands. Science 267:1643–1645.
- 小林朋道. 2007. 水場から離れた高山で見つかったアカハライモリの形態や行動の特異性. 爬虫両棲類学会報. 2007(2) : 120-126
- M.S.Hoogmoed. 1978. An Annoted Review of the Salamander Types Described in the Fauna Japonica. Zoological Mededelingen, 53(9) 98-99
- Matsui K, Marunouchi J, Nakamura M. 2002. An ultrastructural and carotenoid analysis of the red ventrum of the Japanese newt, Cynops pyrrhogaster. Pigment Cell Research 15: 265–272.
- Matsui, K., Mochida, K., & Nakamura, M. (2003). Food habit of the juvenile of the Japanese newt Cynops pyrrhogaster. Zoological science, 20(7), 855-860.
- Mochida, K. 2009. A parallel geographical mosaic of morphological and behavioural aposematic traits of the newt, Cynops pyrrhogaster (Urodela: Salamandridae). Biological journal of the Linnean Society, 97(3), 613-622.
- Mochida, K. 2010. Temperature-dependent aposematic behavior in the newt Cynops pyrrhogaster. Zoological science, 27(7), 555-559.
- Nakada, T., Toyoda, F., Matsuda, K., Nakakura, T., Hasunuma, I., Yamamoto, K., Onoue, S.,& Kikuyama, S. (2017). Imorin: a sexual attractiveness pheromone in female red-bellied newts (Cynops pyrrhogaster). Scientific reports, 7, 41334.
- 佐藤井岐雄. 1977. 日本産有尾類総説. 第一書房, 東京. P358-377
- 沢田昭三. 1961. イモリにおける性行動の地域的差異. 動物学雑誌. 70: 342-347.
- 関慎太朗. 2018. 野外観察のための日本産両生類図鑑. 緑書房, 東京. p.142-143.
- 島田知彦・芹沢俊介. 2013. 見てわかる生物多様性③ たんぼのカエル. 愛知教育大学自然科学系生物領域, 愛知. p25
- Shimada, T., Maeda, S., and Sakakibara, S. 2016. A Morphological Study of cynops phyrrhogaster from the Chita Peninsula :Rediscovery of the “Extinct” Atsumi Race Endemic to PeninsularRgions of Aichi Prefecture,Central Japan. The Hapetological Society of Japan.
- Tagami, M., Horie, C., Kawai, T., Sakabe, A., & Shimada, T. (2015). The mating behavior of Cynops pyrrhogaster from Gifu and Aichi Prefectures, central Japan, in captivity. Current Herpetology, 34(1), 12-18.
- 高谷榮一,大谷勉. 2011. 原色爬虫類両生類検索図鑑. 北陸館, 東京. 220-221p.
- Tominaga, A., Matsui, M., Yoshikawa, N., Nishikawa, K., Hayashi, T., Misawa, Y., & Ota, H. (2013). Phylogeny and historical demography of Cynops pyrrhogaster (Amphibia: Urodela): taxonomic relationships and distributional changes associated with climatic oscillations. Molecular Phylogenetics and Evolution, 66(3), 654-667.
- Tominaga, A., M. Matsui, and Y. Kokuryo. 2015. Occurrence and evolutionary history of two Cynops pyrrhogaster lineages on the Izu Peninsula. Current Herpetology 34:19–27.
- Tominaga, A., Matsui, M., Yoshikawa, N., Eto, K., & Nishikawa, K. 2018 Genome wide displacement and shift of the hybrid zone in the Japanese fire-bellied newt. Journal of Heredity. , 109(3), 232–242
- Tominaga, A., Meyer-Rochow, V. B., Okamoto, T., Kuriyama, T., Nishikawa, K., & Matsui, M. (2016). Origin and Genetic Uniformity of Introduced Population of Cynops pyrrhogaster (Amphibia: Urodela) on Hachijojima Island. Current Herpetology, 35(1), 64-68.